Parallel Stem Cell Niche in Bone Marrow and Oral Cavity – Key to Extramedullary Hematopoiesis and Support of Hematological Malignancies on Oral Tissues
Darling Rojas1, 2, †, Adam Eljarrah1, †, Gagandeep Singh2, †, Lauren S. Sherman2 and Ioanna Tsolaki1, 2,*
1Rutgers School of Dental Medicine, Newark, NJ, USA
2Department of Medicine, Rutgers New Jersey Medical School, Newark, NJ, USA
E-mail: ioanna.tsolaki@rutgers.edu
*Corresponding Author
†Equal contribution
Received 02 July 2024; Accepted 10 July 2024
Abstract
This review article addresses an area of the literature with unanswered questions; although relevant to future treatment of leukemias. The literature describes bone marrow stromal microenvironment as support of leukemias. A key stromal support is mesenchymal stem cells (MSCs), which are similar to those identified in oral tissues, including the dental pulp (DPSC). Hematopoiesis has been reported in oral tissues, although the described activity seems to be distinct from extramedullary hematopoiesis at other sites when there is bone marrow dysfunction such as myelofibrosis. Another parallel between the bone marrow and oral tissues is the survival of leukemia cells. This review describes cases in which the dentist diagnoses leukemia, in particular acute myeloid leukemia (AML). The literature showed AML and other hematological malignancies in gingiva, leaving the question of the source of leukemia. We propose that this question is relevant considering the similar stromal support in bone marrow and oral tissues. It is difficult to determine if current treatments can target leukemia in oral tissues. This article consists of information to argue for continued research in this field.
Keywords: Stem cells, dental pulp, bone marrow, hematopoiesis, hematological malignancy, oral cavity, cancer.
Hematopoietic Activity in the Oral Cavity
Hematopoietic activity in healthy adults occurs in the bone marrow [1]. Pathological conditions or inflammation that disrupts bone marrow activity could lead to hematopoietic activity in other organs, referred as extramedullary hematopoiesis [2]. The oral cavity is prone to inflammation, trauma, and disease, raising the question of whether the gingiva and pulp chamber can support extramedullary hematopoiesis. Indeed, bone marrow stem cells have been shown to have tissue regenerative properties, which could include secretome produced from bone marrow stem cells [3]. Due to the identification of similar types of stem cells in the oral cavity, studies indicated that the latter stem cells have potential in regenerative medicine [4]. Interestingly, transplantation of bone marrow-derived mesenchymal stem cells (MSCs) to gingiva led to differentiation of cells similar to dentin-producing odontoblasts, revival of the pulp of damaged teeth, and regenerate periodontal tissue [5–7].
Central to hematopoietic activity are the hematopoietic stem cells (HSCs) that maintain the production of blood and immune cells [1]. The role of HSCs in hematopoiesis and their potential for regeneration within the setting of the oral cavity has been recently investigated. Transgenic mouse models demonstrated the potential for transplanted HSCs to differentiate into cells endogenous to the periodontium and pulpal chamber [8]. Hematopoietic stem progenitor cells (HSPCs), native to the gingiva, are also capable of differentiating into myeloid progenitors within healthy non-inflamed gingiva, indicating hematopoietic activity for the purpose of achieving gingival blood homeostasis [9].
Myeloid and lymphoid progenitor cells have crucial roles in maintaining gingival health with increased activity during the progression of periodontal disease [9, 10]. This occurs partly by the ability of progenitors to differentiate and contribute to inflammation of periodontal disease. Thus, it is likely that the gingiva could be a site of extramedullary hematopoiesis. Since inflammation can influence cellular production and aging, this should be an interesting avenue to determine how the oral cavity changes in the elderly. These studies could change the landscape of treating age-linked gingival conditions.
Increased stromal derived factor-1 (SDF-1) at sites of tissue damage could chemoattract other cells such as bone marrow-derived MSCs [11]. To this end, the next section discusses MSCs that are endogenous to the dental pulp. Since SDF-1 also acts as a chemoattractant for HSCs, the link between an inflammatory environment such as the oral cavity and stem cells need to be considered to gain insights into hematopoietic dysfunction [12, 13]. An understanding of dental pulp stem cells as part of the oral niche in hematopoietic regulation led us to include a brief review of the bone marrow niche.
Bone Marrow Niche – General
The bone marrow harbors hematopoietic and non-hematopoietic cells, such as HSCs, MSCs, endothelial cells, osteoblasts, and osteoclasts. The MSCs give rise to stromal cells including adipocytes, fibroblasts, chondrocytes, and myocytes. In the bone marrow there is a complex interaction between the HSCs and progenitor cells along with supporting cells to regulate hematopoiesis.
The bone marrow has distinct niches with specific supporting cells that affect the function of HSCs. These niches are the osteoblastic and the endosteal niche. The latter comprises osteoblasts and stromal cells that are located in close proximity to the bone, vascular niche, and endothelial cells within vascular structures. As stated, these niches within the bone marrow contain supporting cells unique to that niche. This results in the production of soluble factors that interact with HSCs and their progenitors. HSCs are retained in the bone marrow by adhering to endothelial cells.
Bone Marrow Niche – MSC and Hematopoietic Regulation
MSCs are heterogeneous multipotent cells that show multilineage differentiation such as osteogenic, adipogenic, and chondrocytic cells [14]. MSCs can exert both pro- and anti-inflammatory immune responses through secreted soluble and insoluble factors, as well as contact-dependent mechanisms [14]. MSCs are major cells of the bone marrow niche that regulate hematopoiesis, partly through the production of cytokines [15, 16].
Nestin+ perivascular cells regulate hematopoiesis as evident by their depletion, resulting in reduced HSCs in bone marrow, increased number of HSCs at extramedullary sites, and disruption of HSC homing to the bone marrow [17]. Nestin+ MSCs were able to retain HSCs in the bone marrow partly through the secretion of CXCL12 and SCF [18].
Nestin+ cells are classified as two distinct populations - those that co-express increased NG2 with colocalization within the arteriolar vasculature, and those with dim leptin receptor (Lep-R) that are associated with sinusoids [19]. Interestingly, the differential spatial distribution of Nestin+ MSC subsets can be paired with functional heterogeneity between NG2+ and LepR+ cells [18]. Depletion of CXCL12 from NG2+/Nestin+ arteriolar cells, but not Nestin-/LepR+ sinusoidal cells, caused HSC cycling, reduced HSC frequencies in the bone marrow, and further resulted in HSC mobilization into peripheral blood and extramedullary sites [18]. In contrast, SCF depletion from low Nestin +/LepR+ cells, but not Nestin high/NG2+ cells, decreased the number of bone marrow HSCs [18]. The major role of MSCs in hematopoietic regulation is supported by the vascular niche, which is discussed in the next section.
Vascular Endothelial Niche in Bone Marrow and at Extramedullary Sites
Previous studies that combine dye uptake assays, label for hypoxic markers, and functional HSC assays, indicated disparate endosteal and vascular niches within the bone marrow. These studies also suggested that HSCs could be found in less perfused and more hypoxic endosteum, as well as distribution throughout the vasculature [20–23]. Owing to advance imaging techniques with improved spatial resolution, recent studies have modified this model by delineating areas of significant overlap between the vasculature and endosteal region, which itself contains the transition from arteriolar to venous circulation [19, 24–27]. Further, arteriolar and sinusoidal vasculature can be differentiated by morphological, molecular, and functional characteristics [24, 28].
Arteriolar microvessels colocalized with two other regulators involved in HSC maintenance – Nestin+ MSCs and non-myelinating Schwann cells that express high level of CXCL12 and SCF, suggesting intrinsic HSC-supportive functions [17, 19, 28–30]. Conditional deletion of SCF from endothelial cells reduced HSCs within bone marrow, without causing significant changes to hematopoietic activity as denoted in the bone marrow, peripheral blood, and splenic cellularity [31]. Endothelial cell subsets, however, mirror the complexity observed between MSC cell subsets by displaying differential contributions to HSC function. While SCF from arteriolar endothelial cells maintains the HSC pool and supports HSC regeneration after myeloablation, the depletion of SCF in sinusoidal endothelial cells has no effect on HSC frequencies, homing, and reconstitution ability [30]. Thus, SCF derived from arteriolar and sinusoidal endothelial cells produce varying effects on HSC numbers and function [30].
It is important to discuss sinusoidal vasculature in extramedullary sites because an understanding of such change at extramedullary sites could provide information on gingival support of hematopoiesis. The literature described sinusoidal vasculature at extramedullary sites in relation to their association with HSCs [21]. Additionally, endothelial cell-derived CXCL12 and SCF have roles in hematopoiesis at extramedullary sites [32]. In fact, the report indicated that CXCL12/CXCR4 are expressed in human gingival fibroblasts in periodontal disease [33]. This is an interesting report when one considers that fibroblasts are major cells of bone marrow stroma that support hematopoietic activity [34]. The parallels described in this section points to similar hematopoietic activity in the gingiva and perhaps other areas of the oral cavity.
Aligning with prior models describing a hierarchical distribution of HSPCs throughout the BM [35, 36], quiescent HSCs were found to be more likely associated with small arterioles and endosteal region, relevant to cycling HSCs. The latter are more likely to be located within the central sinusoidal vasculature and whose association with arterioles could not be distinguished by random distribution [19, 25]. However, this localization may only apply to a subset of HSCs, as the majority of both dormant and cycling HSCs are found in association with sinusoids [21, 26, 32]. Several factors may explain these seemingly contradictory findings. First, associations between HSCs and various bone marrow populations may reflect the frequency of those niche populations, rather than preferential colocalization [26].
In addition to the above, cytokines derived from vascular niche cells, mediate critical HSC functions with cytokine gradients across cell diameters, and within the dense multicellular microenvironments [30, 34]. Thus, tight associations between cell populations may not be necessary for niche cells to exert their HSC-supportive functions. Finally, the possibility that quiescent and cycling HSCs occupy heterogenous niches within the sinusoidal vasculature has not been ruled out [37]. Indeed, while arteriolar vasculature is reduced in aged BM, sinusoids retain their phenotypic and functional properties during aging, and HSCs with the most reconstituting ability, are perisinusoidal in the aged BM. The lends support to the idea that sinusoidal niches themselves promote HSC function [28, 38, 39]. Further support for this framework can be found in a study demonstrating that granulocyte and dendritic cell-monocyte differentiation occurs in spatially distinct compartments within the sinusoidal vasculature [40].
Dental Pulp Mesenchymal Stem Cells (dpMSC)
The section discusses dpMSCs from the oral cavity for subsequent discussion in hematopoietic activity. Encased within the superficial crown of the tooth lies the dental pulp chamber, serving functions related to sensation, nourishment, and tissue regeneration. The center of the chamber houses the major nerves and microvasculature that nourishes the tooth as well as various other cell types that make up the fibrous pulp tissue. These include fibroblasts, dendrites, macrophages, and dpMSCs [41]. The neurovascular bundle enters and exits the tooth structure through an apical foramen at the root of the tooth. The superior and inferior alveolar arteries are responsible for perfusing the maxilla and mandible, including associated structures like the teeth and the gingiva, illustrating the closely interconnected relationship between pulpal and gingival circulation [42]. The multipotent nature of dpMSCs allows for the cells to differentiate into tissues native to the periodontium and gingiva as well as peripheral adipose and neuronal tissue [43]. Similar to bone marrow-derived MSCs, dpMSCs can also exert regenerative and restorative function that benefit the oral cavity, which is prone to damage caused by inflammation and trauma [44].
Inflammation in the Oral Cavity
Due to the role of the oral cavity as the organ that is linked to the open during eating and communication, it is no wonder that the surrounding tissues are exposed and colonized by multiple bacteria. The immune system responds with ongoing inflammation as a method to regulate the colonized organisms [45]. Extensive studies with salivary samples indicated that Developmental endothelial locus-1 (Del-1), lymphocyte function antigen-1 (LFA-1), and IL-17 levels could serve as biomarkers to assess the clinical health and disease status of patients with periodontitis [46].
The focus on Del-1 is interesting since it is an anti-inflammatory protein with three epidermal growth factor-like repeats at the N-terminus [46]. Del 1 is produced by endothelial cells to regulate inflammation and bone loss. This occurs by Del 1 decreasing IL-17 to recruit neutrophils to the periodontium. Binding of Del-1 to 2 integrins blunts the interaction with LFA-1. Del-1-LFA-IL-17 axis is one example of the complex immune interaction in the oral cavity. Cytokines and other inflammatory mediators, together with other factors, show a complex network that maintains oral cavity homeostasis. Table 1 shows examples of cytokines and chemokines in the gingiva and their link to periodontitis.
Table 1 Inflammatory mediators in the gingival disease
| Tissue Insult | Inflamed Mediators | Remarks | Ref. |
| Inflammation Smoking Stress on gingival crevicular |
Elevated IL-1, IL-6, IL-8 | IL-4 is inversely linked to periodontal status – Increased in healthy group | [47] |
| Inflamed Gingival Tissue | High IL-6 and iNOS expression | [48] | |
| Proinflammatory Cytokines: Gingiva vs. Adjacent Sulcular Depth | IL-8IL-6 IL-8 IL-1- | 6 mm 6 mm | [49] |
| Salivary cytokine levels in early gingival inflammation | Decreased IL-1, IL-1Ra, and VEGF | Resolved with time | [50] |
| Cytokines in inflamed gingival tissue and serum samples – Chronic periodontitis | IL-1, TNF-, IL-2, IFN- high in both samples. | [51] |
Leukemia – Bone Marrow and Gingiva
Leukemias are a group of malignant disorders, characterized as acute myeloid leukemia (AML), acute lymphoblastic leukemia (ALL), chronic myelogenous leukemia (CML), and chronic lymphocytic leukemia (CLL). Their etiology is unknown and although termed leukemia, each differs with regards to biology, epidemiology, clinical course, and prognosis. However, most of the leukemias show abnormalities in chromosomes and gene expression which shift normal gene expression or function. This contributes to the leukemic transformation. Many studies have shown that the leukemic cells interaction with the bone marrow stromal cell microenvironment plays a significant role in their survival [52].
Leukemia in Oral Cavity
Systemic diseases such as leukemia show oral changes which may be the first and only signs of disease in patients [53]. Leukemic patients commonly exhibit manifestations in the oral cavity that include petechiae or spontaneous bleeding, mucosal ulceration, and gingival enlargement, with or without necrosis [53, 54]. These are the common initial diagnostic oral manifestations in leukemic patients. A frequent and early finding in leukemic patients is oral lesions, especially at the acute stage of the disease. Myeloid and monocytic/monoblastic leukemia are the more common malignancies showing the discussed oral manifestations [53, 54].
A systematic review described key studies on oral leukemias [55]. This report descrines on on 152 cases of various types of leukemia presenting with oral lesions with 82 cases showed that the most frequent signs were bleeding from the gums, swelling, necrosis, and ulceration of the gingivae. In another report describes frequent occurrences in AML include acute swelling of the gingivae with the gums submerging the teeth frequently occurs. This report also stated that infiltrative lesion of the gum is a strong indication of AML. In 5% of AML, gingival infiltration is the initial manifestation. This is seen more frequently in myelomonocytic (M4) and monocytic (M5) leukemia. When discussing the involvement of thegingiva in leukemia, it was hypothesized that the microanatomy and the expression of endothelial adhesion molecules allows for the infiltration of leukocytes, leading to an overgrowth that is soft in consistency [53].
There are various causes of gingival enlargement and ulceration. It can be caused by neutropenia or a direct infiltration of the immature (blasts) proliferating leukocytes. It can also be secondary to thrombocytopenia and immunodeficiency [56]. The infiltration can lead to increased thickness in the gingiva. This increased thickness results in periodontal pseudo-pockets to mediate secondary inflammatory infiltration [56]. The ongoing movement of myeloid cells in the specialized post-capillary venules leads to the exit of these cells from the circulation and into the gingiva tissue to induce gingivitis and periodontitis.
It is important to note that edentulous individuals (lacking teeth) do not show leukemic infiltration of the gingiva. This could suggest that pathogenesis is associated with local irritation or trauma of the teeth [57]. In one study, cases of metastatic lesions to the oral mucosa, examining literature searches from 1916 to 2011, resulted in 207 cases [54]. Among these, 156 cases reported information on the relevance of teeth. The authors found that 138 showed metastases in dentulous individuals as compared to 18 in edentulous individuals. It is also important to note that in dentulous individuals most of the metastases were found in the gingiva whereas in edentulous individuals these metastases were distributed to other mucosal sites mainly the tongue. It was also evident that 125 cases showed the gingiva was the most common site of metastases regardless of sex. In most of the cases, the alveolar bone was not involved in gingival lesions. Both the maxilla and the mandible were also equally affected [58]. This was a fascinating report that indicated the need to examine the oral cavity when treating malignancies such as leukemia.
CXCL12-CXCR4 Axis in Oral Hematopoiesis
The key role of CXCL12-CXCR4 discussed above led us to include a discussion on this axis in hematopoietic activity in the oral tissue. Bone marrow stroma, which also include endothelial cells produce CXCL12, and constitutes a complex regulatory networks to retain HSCs in the bone marrow to regulate hematopoiesis [52]. Interaction between CXCL12 and CXCR4 is pivotal to hematopoietic regulation, as well as roles in hematologic malignancies [52]. This complexity was demonstrated in studies showing CXCR4 positive HSCs converting into quiescent cells in the presence of CXCL12, and CXCR4 negative HSCs proliferated with CXCL12 [52]. This finding is important to extrapolate to dysfunction of hematological malignancies.
CXCR4 is a member of the CXC chemokine receptor family with seven-transmembrane domains, and activates GTP-binding proteins. CXCR4 is expressed on peripheral blood cells such as neutrophils and monocytes, and HSCs. The chemokine, CXCL12, promotes the migration of peripheral blood cells expressing CXCR4, including pro-B and pre-B cells. CXCR4 activation results in its phosphorylation and endocytosis through clathrin-coated pits. Endocytosed CXCR4 could be ubiquitinated, or recycled to the cell surface [59]. CXCR4 activation is needed to facilitate cells to be chemoattracted towards tissue CXCL12 [60].
Myeloid and lymphoid leukemia cells express CXCR4. Interaction between CXCL12 and CXCR4 on HSC and other bone marrow cells can retain leukemia cells in the bone marrow. CXCR4 is highly expressed in acute leukemias with acute promyelocytic leukemia (subtype M3), myelomonocytic (AML), subtypes M4 and M5, and B-lineage acute lymphocytic leukemia (ALL). The density of membrane CXCR4 correlates to pseudoemperipolesis, which is the migration of cells beneath a layer of bone marrow stromal cells [61].
Variable expression of CXCR4 on AML blasts could be functionally relevant to pseudoemperipolesis [62]. However, in CLL, pseudoemperipolesis is mediated by CXCR4 and CXCL12 [62]. Pseudoemperipolesis is not limited to malignant cells, since similar processes have been reported for normal hematopoietic cells, including lymphocytes, HPCs and HSCs [63]. This is in line with a supporting role for stromal cells in hematopoietic regulation as well as retention of HSCs in the bone marrow [15]. Thus, it is plausible that malignant cells act in a similar manner to survive.
CXCL12 and CXCR4 can promote leukemia progression through other intracellular pathways. PI3K/Akt and MAPK pathways are activated by the binding of CXCL12 to CXCR4 to promote the survival and proliferation of leukemia cells [64]. CXCL12 activates the NF-B pathway to promote the production of soluble factors. These include matrix metalloproteinases, IL-8 and VEGF. Together, these genes facilitate degradation of the extracellular matrix and promote blood vessel formation. It appears that CXCL12 induces increases in the affinity of integrins (VLA-4 and VLA-5) to increase the affinity of blasts to particular niches in bone marrow microenvironment [65]. Interestingly, the infiltration of non-hematopoietic tissues such as gingiva or skin is most often seen with high CXCR4-expressing AML (monocytic differentiation or FAB M4 and M5) [52]. We deduce that that CXCL12-induced migration could be important for the survival of leukemia cells at extramedullary sites such as the oral cavity.
The general thoughts by the scientific and clinical community is that the oral tissue is not the preferred site for primary hematological tumors. Rather, it is believed that metastases to the oral tissues are the result of secondary metastatic sites. A study showed that the most common primary site was the lungs, followed by the kidney, skin, and breasts. The primary site was also sex dependent – for males, the primary sites were the lung, followed by the kidney (14.4%), and skin; females, the primary sites were the breast, followed by genital organs, and lung [66]. Although metastases to the oral mucosa are uncommon, when the malignant cells reach the oral mucosa, the favored site is the gingiva. Due to the gingiva being susceptible to inflammation, it is believed that inflammatory mediators could be one of the reasons why many cancers show preference for the gingiva. The literature reported 42 of the 207 cases of oral cancer were found with gingival inflammation. Of the 42 cases, 38 were shown in the area of gingival inflammation [66]. One reported hypothesis for the role of gingival hyperplasia pertaining to leukemia is that the periodontal bacteria present low-level antigens that act as chemoattractants for myelomonocytic leukemic cells [67]. Tumor associated macrophages may also be attracted or induced by oral cytokines [68].
In rare cases, the evidence suggested that cancer could originate from the oral cavity and metastasize to other tissues. Here we examine examples of the oral tissue as the primary or secondary site of metastasis [69]. The most common tumor affecting the oral cavity is oral squamous cell carcinoma (SCC). This tumor type accounts for about 90% of all oral cancers [69, 70]. SCC does not normally affect the gingiva, but 10% of cases have been reported to be in the gingiva. SCC generally affects the lateral ventral surface of the tongue, the floor of the mouth and buccal mucosa. There is some uncertainty as to whether SCC affects males and females equally as it has been reported to have a predominance in both sexes.
Unlike leukemia, oral cancer seems to favor edentulous areas compared to dentate areas. It is also more common to arise in the mandibular gingiva compared to the maxillary gingiva. It also seems to favor the gingiva posterior to the premolars in 60% of the cases. These carcinomas can go undetected as they are misdiagnosed as benign tumors or gingival inflammation. There is a case report of a 62-year-old female patient who visited a dental clinic complaining of pain in the right posterior area of the mandible. An oral examination revealed that she had a buccal gingival growth near tooth #47 that was reddish in color which measured 0.5 cm 0.5 cm. Upon further examination the rest of her dentations showed signs of generalized chronic periodontitis. Initially the growth was diagnosed as apical periodontitis and inflammatory/reactive gingival growth in relation to tooth #47. The patient was prescribed antibiotics, analgesics, and chlorhexidine mouthwash for a duration of 3 days and released. She was told to report back in a week. When the patient returned, there was no improvement with respect to growth. The patient requested to have the tooth removed, which was respected. The dentist suspected oral squamous cell carcinoma and metastatic carcinoma to the gingiva and referred her to the appropriate specialist. A biopsy and radiograph showed a soft tissue mass growing in the extraction site very rapidly. The mass was measuring about 1 cm 1 cm from the site of extraction. During histopathological examination it was shown that islands and sheets of dysplastic epithelium invaded the underlying connective tissue stroma along with keratin pearl formation. Additionally, the overlying epithelium exhibited hyperkeratinized stratified squamous epithelium with dysplastic features indicating a diagnosis of well-differentiated SCC. The literature suggested that curettage or extraction of the tooth could worsen the prognosis as the invasive procedure could increase the risk of metastases to distant sites by spreading the cancer cells into circulation [71].
Table 2 Representative landscape of cancers in gingiva
| Cancer | Diagnosis Site | Age/Sex | Ref. |
| AML | Gingiva | 18 yrs/Female | [53] |
| AML* | Gingiva | 34 yrs/Female | [56] |
| Oral SCC | Gingiva | 49 yrs/Male | [69] |
| Myeloid sarcoma ** | Gingiva | 24 yrs/Female | [73] |
| AML (subtype M4) | Gingiva | 10 yrs/Female | [74] |
| AML | Gingiva | 23 yrs/Female | [75] |
| *FAB classification M4 | |||
| **Associated with acute promyelocytic leukemia | |||
Another case of a cancers that originate in the oral cavity is described in this paragraph [69]. In a case of malignant melanoma that originated in the maxillary gingiva, a 54-year-old man with tooth extraction 4 months was presented with a growth in the upper left maxillary gingiva. It had been present for the past 4 months and recently started bleeding in the past 15 days. It was also noted that he had a spot adjacent to the extracted tooth that was black/ bluish in color and was soft in consistency, ulcerated and nontender. A biopsy was performed, and he was diagnosed with malignant melanoma. The patient refused further treatment. It should be noted that primary melanomas of the oral cavity are rare. The most common sites affected in the oral cavity by melanomas are the palate and maxillary gingiva. These are then followed by the mandibular gingiva, the buccal mucosa, the tongue and the floor of the mouth [72].
In a rare case report of SCC with the primary site being the mandibular gingiva, metastasized to the small intestine. This was rare since cancers originating from the head/neck have not been reported to metastasize to the small intestine. The 82-year-old male visited a clinic complaining of swelling and pain in his right mandibular gingiva and the right submandibular region for 4 months. A physical examination and imaging revealed a 25 mm-sized exophytic tumor in the right mandibular gingiva, and metastasis was confined to the cortical bone of the mandible. Computed tomography (CT) and Positron emission tomograph (PET) scans showed metastases to the cervical lymph node at the right superior neck and right mid internal jugular section, abnd mandibular gingiva. Following neoadjuvant treatment, secondary tumors in the small intestine was attributed to hematogenous route. Table 2 lists a few cases in which the cancer was identified in the gingiva but is unsure if the tumor originated at the site or if this tumors were results of secondary metastasis.
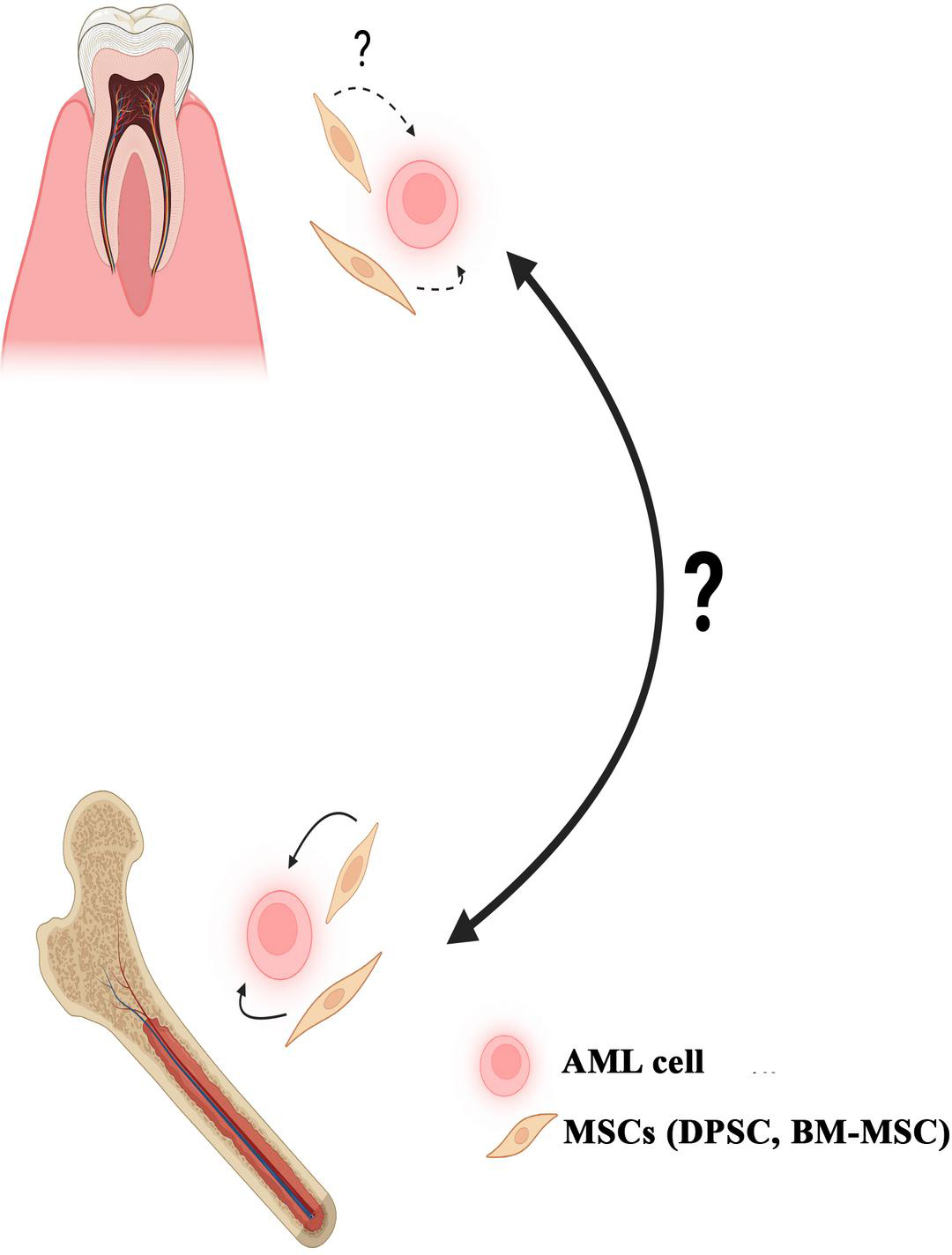
Figure 1 Key points from this review article. Shown are AML cells representing other leukemias that are supported by bone marrow derived mesenchymal (BM-MSCs), and similar oral cells, dental pulp derived mesenchymal stem cells (DPSCs). Shown are similar support by DPSCs. The question mark indicates the uncertainty of leukemia, oral tissue vs. bone marrow.
Figure 1 summarizes the key message in this brief review article. Bone marrow-derived mesenchymal stem cells (BM-MSCs) support AML. Its counterpart in the oral cavity, dental pulp-derived stem cells (DPSCs), may serve a similar function to support AML and other leukemia in the gingiva. Also discussed, is the literature on hematopoietic activity in the oral tissues. Although there are several case reports on AML in the oral tissues, it is unclear where leukemia originates. The general consensus is that leukemia develops in the bone marrow. However, the similarity in cell types, and hematopoietic activity form the basis for the validity of asking this question. This review outlines the need for future studies in experimental models to understand how the bone marrow microenvironment and oral tissues are involved in leukemia development. Another important question, is if the current treatments can equally target leukemia in the bone marrow and gingiva with similar efficiency.
Acknowledgments
This work is in partial fulfillment for a doctoral thesis by IT.
References
[1] Crane, G. M., E. Jeffery, and S. J. Morrison. 2017. Adult haematopoietic stem cell niches. Nat Rev Immunol 17: 573–590.
[2] Mende, N., and E. Laurenti. 2021. Hematopoietic stem and progenitor cells outside the bone marrow: where, when, and why. Exp Hematol 104: 9–16.
[3] Yoshikawa, T., T. Noshi, H. Mitsuno, K. Hattori, K. Ichijima, and Y. Takakura. 2001. Bone and soft tissue regeneration by bone marrow mesenchymal cells. Materials Sci Eng C 17: 19–26.
[4] Kawaguchi, H., A. Hirachi, N. Hasegawa, T. Iwata, H. Hamaguchi, H. Shiba, T. Takata, Y. Kato, and H. Kurihara. 2004. Enhancement of periodontal tissue regeneration by transplantation of bone marrow mesenchymal stem cells. J Periodontol 75: 1281–1287.
[5] Li, Z. Y., L. Chen, L. Liu, Y. F. Lin, S. W. Li, and W. D. Tian. 2007. Odontogenic potential of bone marrow mesenchymal stem cells. J Oral Maxillofac Surg 65: 494–500.
[6] Ishizaka, R., K. Iohara, M. Murakami, O. Fukuta, and M. Nakashima. 2012. Regeneration of dental pulp following pulpectomy by fractionated stem/progenitor cells from bone marrow and adipose tissue. Biomaterials 33: 2109–2118.
[7] Kawaguchi, H., A. Hirachi, N. Hasegawa, T. Iwata, H. Hamaguchi, H. Shiba, T. Takata, Y. Kato, and H. Kurihara. 2004. Enhancement of periodontal tissue regeneration by transplantation of bone marrow mesenchymal stem cells. J Periodontol 75: 1281–1287.
[8] Wilson, K. R., I.-H. Kang, U. Baliga, Y. Xiong, S. Chatterjee, E. Moore, B. Parthiban, K. Thyagarajan, J. L. Borke, and S. Mehrotra. 2018. Hematopoietic stem cells as a novel source of dental tissue cells. Sci Reports 8: 8026.
[9] Krishnan, S., K. Wemyss, I. E. Prise, F. A. McClure, C. O’boyle, H. M. Bridgeman, T. N. Shaw, J. R. Grainger, and J. E. Konkel. 2021. Hematopoietic stem and progenitor cells are present in healthy gingiva tissue. J Exp Med 5: 218.
[10] Hajishengallis, G., and J. M. Korostoff. 2017. Revisiting the Page & Schroeder model: the good, the bad and the unknowns in the periodontal host response 40 years later. Periodontol 2000 75: 116–151.
[11] Kimura, Y., M. Komaki, K. Iwasaki, M. Sata, Y. Izumi, and I. Morita. 2014. Recruitment of bone marrow-derived cells to periodontal tissue defects. Frontiers Cell Dev Biol 2: 19.
[12] Hattori, K., B. Heissig, and S. Rafii. 2003. The regulation of hematopoietic stem cell and progenitor mobilization by chemokine SDF-1. Leuk lymphoma 44: 575–582.
[13] Mendt, M., and J. E. Cardier. 2015. Role of SDF-1 (CXCL12) in regulating hematopoietic stem and progenitor cells traffic into the liver during extramedullary hematopoiesis induced by G-CSF, AMD3100 and PHZ. Cytokine 76: 214–221.
[14] Sherman, L. S., A. Condé-Green, O. A. Sandiford, and P. Rameshwar. 2015. A discussion on adult mesenchymal stem cells for drug delivery: pros and cons. Ther Deliv 6: 1335–1346.
[15] Ehninger, A., and A. Trumpp. 2011. The bone marrow stem cell niche grows up: mesenchymal stem cells and macrophages move in. J Exp Med 208: 421–428.
[16] Aqmasheh, S., P. Akbarzadehlaleh, D. P. Sarvar, and H. Timari. 2017. Effects of mesenchymal stem cell derivatives on hematopoiesis and hematopoietic stem cells. Adv Pharmaceutical Bulletin 7: 165.
[17] Méndez-Ferrer, S., T. V. Michurina, F. Ferraro, A. R. Mazloom, B. D. MacArthur, S. A. Lira, D. T. Scadden, A. Ma’ayan, G. N. Enikolopov, and P. S. Frenette. 2010. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 466: 829–834.
[18] Asada, N., Y. Kunisaki, H. Pierce, Z. Wang, N. F. Fernandez, A. Birbrair, A. Ma’ayan, and P. S. Frenette. 2017. Differential cytokine contributions of perivascular haematopoietic stem cell niches. Nature Cell Biol 19: 214–223.
[19] Kunisaki, Y., I. Bruns, C. Scheiermann, J. Ahmed, S. Pinho, D. Zhang, T. Mizoguchi, Q. Wei, D. Lucas, and K. Ito. 2013. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 502: 637–643.
[20] Eliasson, P., and J. I. Jönsson. 2010. The hematopoietic stem cell niche: low in oxygen but a nice place to be. J Cell Physiol 222: 17–22.
[21] Kiel, M. J., Ö. H. Yilmaz, T. Iwashita, O. H. Yilmaz, C. Terhorst, and S. J. Morrison. 2005. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell 121: 1109–1121.
[22] Parmar, K., P. Mauch, J.-A. Vergilio, R. Sackstein, and J. D. Down. 2007. Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia. Proc Natl Acad Sci 104: 5431–5436.
[23] Winkler, I. G., N. A. Sims, A. R. Pettit, V. Barbier, B. Nowlan, F. Helwani, I. J. Poulton, N. van Rooijen, K. A. Alexander, and L. J. Raggatt. 2010. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 116: 4815–4828.
[24] Kusumbe, A. P., S. K. Ramasamy, and R. H. Adams. 2014. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 507: 323–328.
[25] Nombela-Arrieta, C., G. Pivarnik, B. Winkel, K. J. Canty, B. Harley, J. E. Mahoney, S.-Y. Park, J. Lu, A. Protopopov, and L. E. Silberstein. 2013. Quantitative imaging of haematopoietic stem and progenitor cell localization and hypoxic status in the bone marrow microenvironment. Nature Cell Biol 15: 533–543.
[26] Christodoulou, C., J. A. Spencer, S.-C. A. Yeh, R. Turcotte, K. D. Kokkaliaris, R. Panero, A. Ramos, G. Guo, N. Seyedhassantehrani, and T. V. Esipova. 2020. Live-animal imaging of native haematopoietic stem and progenitor cells. Nature 578: 278–283.
[27] Kokkaliaris, K. D., L. Kunz, N. Cabezas-Wallscheid, C. Christodoulou, S. Renders, F. Camargo, A. Trumpp, D. T. Scadden, and T. Schroeder. 2020. Adult blood stem cell localization reflects the abundance of reported bone marrow niche cell types and their combinations. Blood 136: 2296–2307.
[28] Kusumbe, A. P., S. K. Ramasamy, T. Itkin, M. A. Mäe, U. H. Langen, C. Betsholtz, T. Lapidot, and R. H. Adams. 2016. Age-dependent modulation of vascular niches for haematopoietic stem cells. Nature 532: 380–384.
[29] Yamazaki, S., H. Ema, G. Karlsson, T. Yamaguchi, H. Miyoshi, S. Shioda, M. M. Taketo, S. Karlsson, A. Iwama, and H. Nakauchi. 2011. Nonmyelinating Schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 147: 1146–1158.
[30] Xu, J., Y. Wang, P. Guttorp, and J. L. Abkowitz. 2018. Visualizing hematopoiesis as a stochastic process. Blood Adv 2: 2637–2645.
[31] Ding, L., T. L. Saunders, G. Enikolopov, and S. J. Morrison. 2012. Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 481: 457–462.
[32] Inra, C. N., B. O. Zhou, M. Acar, M. M. Murphy, J. Richardson, Z. Zhao, and S. J. Morrison. 2015. A perisinusoidal niche for extramedullary haematopoiesis in the spleen. Nature 527: 466–471.
[33] Hosokawa, Y., I. Hosokawa, K. Ozaki, H. Nakae, K. Murakami, Y. Miyake, and T. Matsuo. 2005. CXCL12 and CXCR4 expression by human gingival fibroblasts in periodontal disease. Clin Exp Immunol 141: 467–474.
[34] Ogawa, M., A. C. LaRue, and C. J. Drake. 2006. Hematopoietic origin of fibroblasts/myofibroblasts: Its pathophysiologic implications. Blood 108: 2893–2896.
[35] Dzierzak, E., and N. A. Speck. 2008. Of lineage and legacy: the development of mammalian hematopoietic stem cells. Nature Immunol 9: 129–136.
[36] Doulatov, S., F. Notta, E. Laurenti, and J. E. Dick. 2012. Hematopoiesis: a human perspective. Cell Stem Cell 10: 120–136.
[37] Acar, M., K. S. Kocherlakota, M. M. Murphy, J. G. Peyer, H. Oguro, C. N. Inra, C. Jaiyeola, Z. Zhao, K. Luby-Phelps, and S. J. Morrison. 2015. Deep imaging of bone marrow shows non-dividing stem cells are mainly perisinusoidal. Nature 526: 126–130.
[38] Kusumbe, A. P. 2016. Vascular niches for disseminated tumour cells in bone. J Bone Oncol 5: 112–116.
[39] Saçma, M., J. Pospiech, R. Bogeska, W. de Back, J.-P. Mallm, V. Sakk, K. Soller, G. Marka, A. Vollmer, and R. Karns. 2019. Haematopoietic stem cells in perisinusoidal niches are protected from ageing. Nature Cell Biol 21: 1309–1320.
[40] Zhang, J., Q. Wu, C. B. Johnson, G. Pham, J. M. Kinder, A. Olsson, A. Slaughter, M. May, B. Weinhaus, and A. D’Alessandro. 2021. In situ mapping identifies distinct vascular niches for myelopoiesis. Nature 590: 457–462.
[41] Nanci, A. 2008. Enamel: composition, formation, and structure. Ten Cate’s Oral histology: Development, structure, and function. 183–184.
[42] Gopal, S., K. P. Shetty, V. Jindal, and M. Saritha. 2011. Interrelationship of endodontic-Periodonctic-Periodontal lesions- an overview. Indian J Dental Sci 3.
[43] Zhang, W., X. F. Walboomers, S. Shi, M. Fan, and J. A. Jansen. 2006. Multilineage Differentiation Potential of Stem Cells Derived from Human Dental Pulp after Cryopreservation. Tissue Eng 12: 2813–2823.
[44] Gendron, R., D. Grenier, and L.-F. Maheu-Robert. 2000. The oral cavity as a reservoir of bacterial pathogens for focal infections. Microbes Inf 2: 897–906.
[45] Silva, L. 2015. A literature review of inflammation and its relationship with the oral cavity. Global J Inf Dis Clin Res 1: 1–7.
[46] Inönü, E., S. A. Kayis, M. A. Eskan, and S. S. Hakki. 2020. Salivary Del-1, IL-17, and LFA-1 levels in periodontal health and disease. J Periodontal Res 55: 511–518.
[47] Giannopoulou, C., J. J. Kamma, and A. Mombelli. 2003. Effect of inflammation, smoking and stress on gingival crevicular fluid cytokine level. J Clin Periodontol 30: 145–153.
[48] Hirose, M., K. Ishihara, A. Saito, T. Nakagawa, S. Yamada, and K. Okuda. 2001. Expression of Cytokines and Inducible Nitric Oxide Synthase in Inflamed Gingival Tissue. J Periodontol 72: 590–597.
[49] McGee, J. M., M. A. Tucci, T. P. Edmundson, C. L. Serio, and R. B. Johnson. 1998. The Relationship Between Concentrations of Proinflammatory Cytokines Within Gingiva and the Adjacent Sulcular Depth. J Periodontol 69: 865–871.
[50] Belstrøm, D., C. Damgaard, E. Könönen, M. Gürsoy, P. Holmstrup, and U. K. Gürsoy. 2017. Salivary cytokine levels in early gingival inflammation. J Oral Microbiol 9: 1364101.
[51] Górska, R., H. Gregorek, J. Kowalski, A. Laskus-Perendyk, M. Syczewska, and K. Madaliñski. 2003. Relationship between clinical parameters and cytokine profiles in inflamed gingival tissue and serum samples from patients with chronic periodontitis. J Clin Periodontol 30: 1046–1052.
[52] Sison, E. A., and P. Brown. 2011. The bone marrow microenvironment and leukemia: biology and therapeutic targeting. Expert Rev Hematol 4: 271–283.
[53] Hasan, S., N. I. Khan, and L. B. Reddy. 2015. Leukemic gingival enlargement: Report of a rare case with review of literature. Int J Appl Basic Med Res 5: 65–67.
[54] Cammarata-Scalisi, F., K. Girardi, L. Strocchio, P. Merli, A. Garret-Bernardin, A. Galeotti, F. Magliarditi, A. Inserra, and M. Callea. 2020. Oral Manifestations and Complications in Childhood Acute Myeloid Leukemia. Cancers (Basel) 12: 1634.
[55] McCarthy, F. P., and P. H. Karcher. 1946. The Oral Lesions of Monocytic Leukemia. New Engl J Med 234: 787–790.
[56] Menezes, L., and J. R. Rao. 2012. Acute myelomonocytic leukemia presenting with gingival enlargement as the only clinical manifestation. J Indian Soc Periodontol 16: 597–601.
[57] Sheikh, O., and M. Perry. 2021. The Lips, Mouth, Tongue and Teeth: Part II. Diseases and Injuries to the Head, Face and Neck: A Guide to Diagnosis and Management 1085–1168.
[58] Zhang, X., Y. Li, Z. Ge, H. Zhao, L. Miao, and Y. Pan. 2020. The dimension and morphology of alveolar bone at maxillary anterior teeth in periodontitis: a retrospective analysis-using CBCT. Intl J Oral Sci 12: 4.
[59] Marchese, A., and J. L. Benovic. 2001. Agonist-promoted ubiquitination of the G protein-coupled receptor CXCR4 mediates lysosomal sorting. J Biol Chem 276: 45509–45512.
[60] Arya, M., H. Ahmed, N. Silhi, M. Williamson, and H. R. Patel. 2007. Clinical importance and therapeutic implications of the pivotal CXCL12-CXCR4 (chemokine ligand-receptor) interaction in cancer cell migration. Tumor Bio 28: 123–131.
[61] Kim, Y.-R., and K.-S. Eom. 2014. Simultaneous inhibition of CXCR4 and VLA-4 exhibits combinatorial effect in overcoming stroma-mediated chemotherapy resistance in mantle cell lymphoma cells. Immune Network 14: 296–306.
[62] Burger, J. A., A. Spoo, A. Dwenger, M. Burger, and D. Behringer. 2003. CXCR4 chemokine receptors (CD184) and 41 integrins mediate spontaneous migration of human CD34+ progenitors and acute myeloid leukaemia cells beneath marrow stromal cells (pseudoemperipolesis). Br J Haematol 122: 579–589.
[63] Hial, H., Y. Nishi, T. Mlyazawa, Y. Matsudalra, Y. Matsudalra, and Y. Nishizuka. 1981. Mouse lymphoid leukemias: symbiotic complexes of neoplastic lymphocytes and their microenvironments. J Natl Cancer Inst 66: 713–722.
[64] de Lourdes Perim, A., M. K. Amarante, R. L. Guembarovski, C. E. C. de Oliveira, and M. A. E. Watanabe. 2015. CXCL12/CXCR4 axis in the pathogenesis of acute lymphoblastic leukemia (ALL): a possible therapeutic target. Cell Mol Life Sci 72: 1715–1723.
[65] Sison, E. A. R., and P. Brown. 2011. The bone marrow microenvironment and leukemia: biology and therapeutic targeting. Expert Rev Hematol 4: 271–283.
[66] Allon, I., A. Pessing, I. Kaplan, D. M. Allon, and A. Hirshberg. 2014. Metastatic Tumors to the Gingiva and the Presence of Teeth as a Contributing Factor: A Literature Analysis. J Periodontol 85: 132–139.
[67] Bann AlHazmi, B. 2021. Leukemia and periodontal health. J Pakistan Dental Assoc 30(01).
[68] Costa, N. L., M. C. Valadares, P. P. C. Souza, E. F. Mendonça, J. C. Oliveira, T. A. Silva, and A. C. Batista. 2013. Tumor-associated macrophages and the profile of inflammatory cytokines in oral squamous cell carcinoma. Oral Oncol 49: 216–223.
[69] Ramesh, R., and A. Sadasivan. 2017. Oral squamous cell carcinoma masquerading as gingival overgrowth. Eur J Dent 11: 390–394.
[70] Bharanidharan, R., T. Dineshkumar, K. Raghavendhar, and A. R. Kumar. 2015. Squamous cell carcinoma of the gingiva: A diagnostic enigma. J Oral Maxillofac Pathol 19: 267.
[71] Seow, W. K. 1994. Root agenesis associated with chronic infection and traumatic curettage of the maxillary sinus. Pediatr Dent 16: 227.
[72] Pour, M. H. 2008. Malignant melanoma of the oral cavity: a review of literature. Indian Journal of Dental Research 19: 47–51.
[73] de Andrade, B. A., R. B. Farneze, M. Agostini, E. B. Cortezzi, A. C. Abrahão, M. G. Cabral, A. Rumayor, and M. J. Romañach. 2017. Myeloid sarcoma of the oral cavity: A case report and review of 89 cases from the literature. J Clin Exp Dent 9: e1167–e1171.
[74] Fernandes, K. S., M. Gallottini, T. Castro, M. F. Amato, J. S. Lago, and P. H. Braz-Silva. 2018. Gingival leukemic infiltration as the first manifestation of acute myeloid leukemia. Special Care in Dentistry 38: 160–162.
[75] Dantas, R. C. M., V. D. C. Vieira, W. S. MagalhÃEs, M. S. Assis, T. F. L. D. E. Oliveira, V. A. Sarmento, and P. Leite-Ribeiro. 2020. Acte myeloid leukemia with initial manifestation in gingiva: A case report. Oral Surgery, Oral Medicine, Oral Pathol Oral Radiol 130: e219.
International Journal of Translational Science, Vol. 1, 201–220.
doi: 10.13052/ijts2246-8765.2024.034
© 2024 River Publishers